A Connectionist Account of Semantic Impairments and the Access/Degraded-Store Distinction
Stephen J. Gotts and David C. Plaut
Dept. of Psychology, Carnegie Mellon University
& Center for the Neural Basis of Cognition
Introduction
Neuropsychological studies of patients with acquired semantic impairments
have yielded two distinct and contrasting patterns of performance in a
spoken-word/picture matching task (Warrington & Cipolotti,
1996). Patients labeled "access/refractory" are strongly influenced by
presentation rate, semantic relatedness of distractors, and repetition, yet they
seem relatively unaffected by lexical frequency. "Degraded-store" patients, on
the other hand, are strongly affected by lexical frequency but not by presentation
rate, semantic relatedness, or repetition.
In our theoretical account of this pattern of data, language processes maintain
and integrate semantic information over time, thereby compensating for the
natural tendency of cortical neurons to habituate. Damage to parts of the
language system responsible for semantic maintenance/integration has the effect
of leaving semantic representations vulnerable to cortical habituation -
producing the "access/refractory" pattern. Damage to semantic representations
themselves leaves the compensation spared, producing the "degraded-store"
pattern.
Access/Degraded-Store Distinction
Warrington (1975) documented patients with temporal
lobe atrophy sparing medial structures ("semantic dementia") who exhibited selective
semantic deficits. Comprehension performance was better for objects with high-frequency
names.
Warrington & Shallice (1979) documented a patient whose
reading performance was highly inconsistent and unaffected by frequency, yet the
patient could access some semantic information about words he couldn't read.
They proposed criteria to distinguish between patients with impaired semantic
access processes and patients with degraded representations.
Warrington & McCarthy (1983; 1987) studied globally
aphasic patients who seemed to fit with the "access" pattern: Their performance on
a spoken-word/picture matching task was poor under conditions of fast
presentation rate, multiple repetitions of trials and semantically close
distractors.
Rapp & Caramazza (1993) criticized the empirical
validity of the "access/degraded-store" distinction, pointing out that no patient
had been tested with all of the criteria and that a number of "mixed" patterns
had also been observed. They argued that the distinction would be of little
use until more specific theoretical claims were made about the nature of
representations and access procedures.
Warrington & Cipolotti (1996) responded to the
criticism of poor empirical validity, showing that two sets of patients do indeed
present with a contrasting pattern of impairment.
Patients:
Two globally aphasics (A1, A2) with wide-spread damage
of the left hemisphere (more frontal in A1, more posterior
in A2), largely sparing temporal cortex.
Four "semantic dementia" patients (S1-S4), all with focal
atrophy of the left temporal lobe.
Task:
Spoken-word/picture matching (point to a picture in an array
of 4 that matches the word spoken by the experimenter).
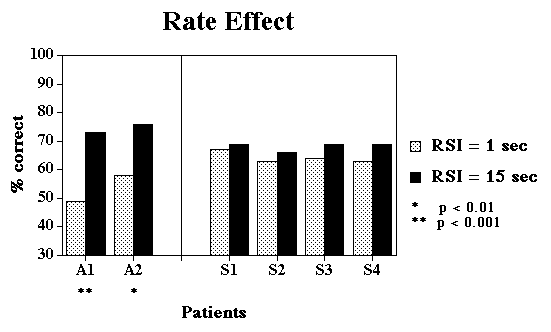
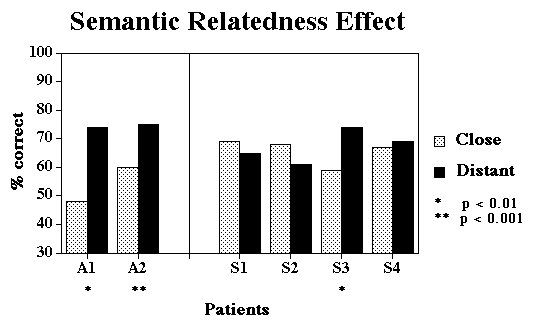
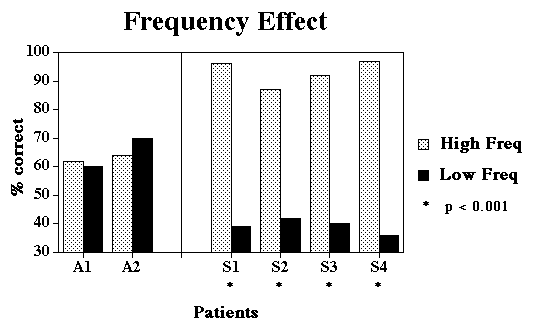
Design:
Response-Stimulus Interval (1 sec vs. 15 secs), Semantic
Relatedness (Close vs. Distant), and Lexical Frequency (High
vs. Low) were completely crossed, and each condition was tested
in a blocked fashion. A block consisted of 3 repetitions of
all 4 pictures in an array, probed in pseudorandom order.

Hypothesis
Language processing requires the maintenance and integration of semantic
information over time. Given numerous observations that neurons in the
brain habituate (i.e. show a response decrement) under conditions of
repeated stimulation, sustaining semantic information should be
particularly difficult.
In order to effectively perform language comprehension and production, we
propose that neurons in temporal cortex which participate in coding semantic
information compensate for habituation by learning to use interactions with
other cortical areas (perhaps frontal and posterior parietal areas).
We suggest that the "access/refractory" patients have damage to areas which
help to sustain and integrate semantic information over time, unmasking the
normal compensation of semantic habituation. For these patients, word/picture
matching performance should be most impaired in the conditions expected to
maximize habituation of semantic information (e.g. fast rate, semantically
close distractors, and multiple repetitions). Frequency should have a
relatively small effect because the factors driving poor performance are
less related to frequency (how many neurons are shared across semantically
similar representations).
We would claim that "degraded-store" patients, on the other hand, have
damage to semantic representations themselves. They shouldn't be affected
as much by rate of presentation and repetition because the cortical areas
compensating for semantic habituation are largely spared. Frequency should
matter more because the neurons coding semantic information provide top-down
support for earlier levels of processing (e.g. phonology). Damage may weaken
the support to the extent that correct performance is affected. Low frequency
words will be most affected because the inputs are particularly weak.
Semantic relatedness might also be expected to matter for these patients,
although if damage is severe enough, a within-category manipulation might
have no effect.
Evidence for Neural Habituation
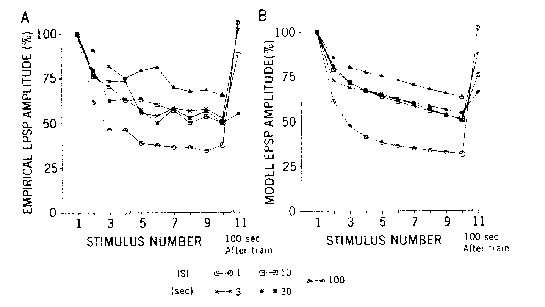
Gingrich & Byrne (1985)
Decremented responses of motor neurons in Aplysia following repeated
stimulation of the sensory neurons at different ISI's (Panel A:
Byrne, 1982). The data are modeled (Panel B) with transmitter depletion and
inactivation of Ca++ channels.
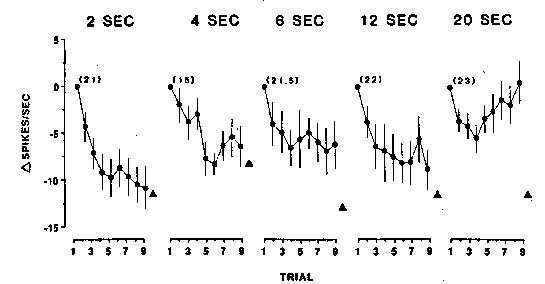
Miller, Gochin, & Gross (1991)
Recordings from neurons in inferior temporal cortex of anesthetized macaque
monkeys during the repeated presentation of common objects at different ISI's.
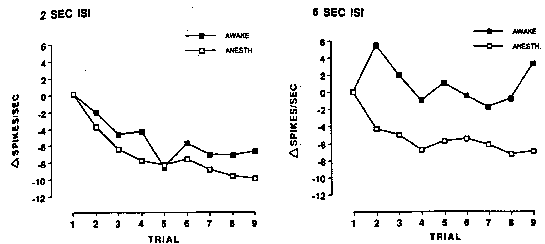
Recordings from anesthetized and awake monkeys performing a passive viewing
task.
Simulation
Rather than modeling spoken-word/picture matching, per se,
we modeled only the auditory word comprehension aspects of
the task. This is partially motivated by the observation
that "access/refractory" patients on record tend to perform at ceiling
on picture/picture matching.
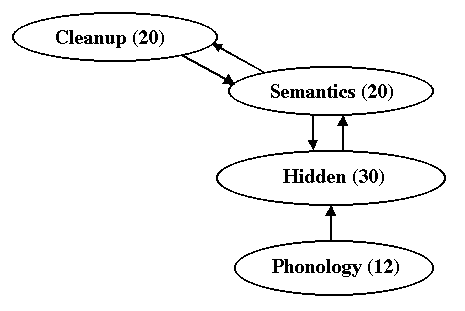
Network Architecture
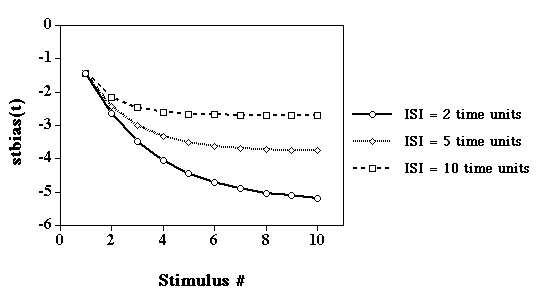
Habituation
Short-term bias added to the netinput of each unit in the network:
stbiasj(t) = -c * aj(t-1) + lambda * stbiasj(t-1)
where c is the "buildup parameter, lambda is the "decay"
parameter, and aj(t-1) is the activation of unit
j at time t-1. The values of c and lambda were
constrained to be the same for all units in the network. (See
Staddon & Higa, 1996, for a similar modeling
approach.)
Training Procedure
Input Patterns:
a "word" is a temporal sequence of 2 phonological
patterns or "phonemes"
8 unique phonemes (4/12 units on for each)
32 total words were constructed from the 8 phonemes.
Target Patterns:
32 semantic patterns (5/20 units on for each) forming
2 non-overlapping categories (16 members each)
In each category, 8 patterns are "closely" related (on
average, share 3.875 "on" units with category prototype),
and 8 are "distantly" related (share 2.375 "on" units with
category prototype).
Each semantic pattern is paired at random with an
input "word".
The network was trained with a continuous, temporal
version of backpropagation discussed in Pearlmutter
(1989).
Half of the 32 training patterns were assigned to be high
frequency (presented twice as often during training) and
half low frequency.
Time Course of a Single Training Pattern:
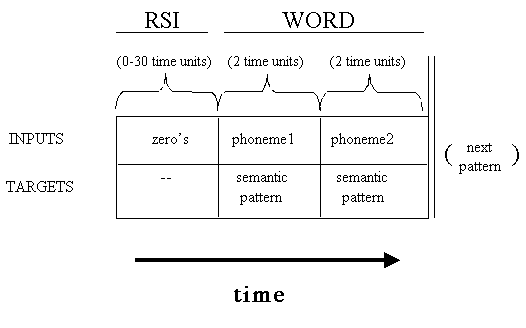
RSI's were biased to be short if members from the same
category followed each other.
Results:
After approximately 30,000 passes through the
training set, the network performed at 91% correct
(on best-match criterion described in "Testing
Procedure").
Lesioning Procedure
"Access/refractory" damage:
randomly removed connections between cleanup
and semantics
"Degraded-Store" damage:
randomly removed semantic units
Damage was sampled to produce a comparable level of
performance to that of the patients and matched
performance for the two types of damage. Each lesion
severity explored was repeated 20 times to insure a
stable estimate of performance.
Testing Procedure
Each lesioned network was presented with 4 types of arrays
of 4 words each, as in Warrington & Cipolotti (1996):
Close/HF, Close/LF, Distant/HF, Distant/LF.
In each testing block, all 4 words in one of the arrays were
probed 3 times in a pseudorandom order and at a fixed
RSI (either 2 or 30 time units).
Each array was presented at both a fast rate and a slow rate.
The pattern of semantic activity generated by an input word
was compared with the target semantic patterns of all the
words in the array. The best match was taken to be the
network's response (chance performance = 25% correct).
Simulation Results
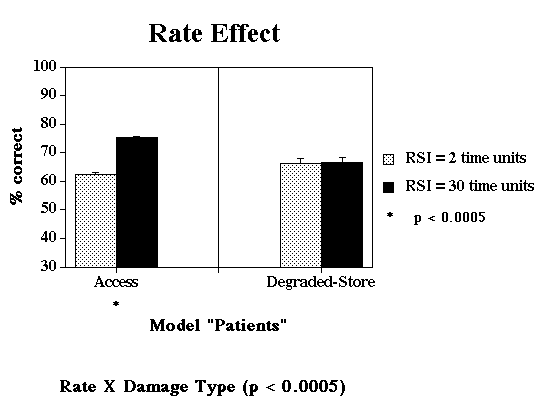
Jump to Patient Data: Rate Effect
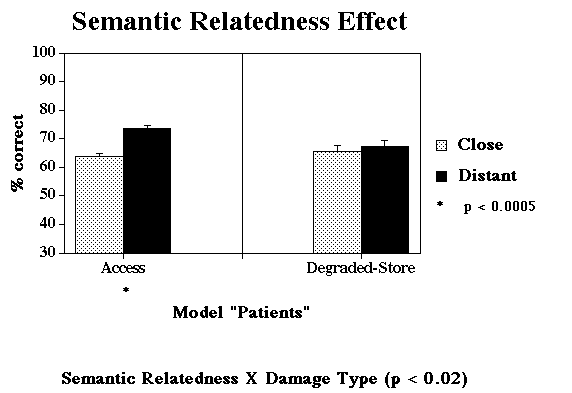
Jump to Patient Data: Semantic Relatedness Effect
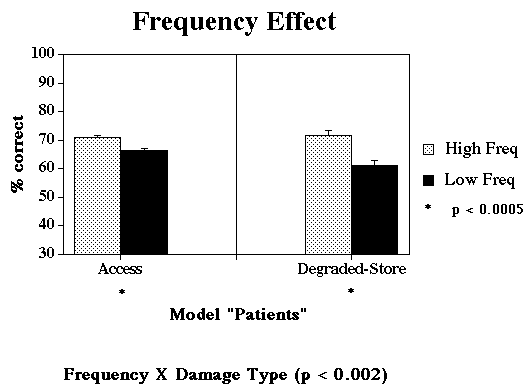
Jump to Patient Data: Frequency Effect

Jump to Patient Data: Consistency & Serial Position Effects
Conclusions
A connectionist model which is trained to maintain and
integrate semantic information and which incorporates
the principle of neural habituation is capable of accounting
for the basic pattern of data associated with the "access/
degraded-store" distinction. While the model is capturing
all of the relevant effects, some of the effects are not
sufficiently strong in magnitude (e.g. the frequency effect,
under "degraded-store" damage). Current modeling
efforts are aimed at providing a better fit to the data.
Because the theorizing relies on neural habituation as a
principle of the normal cognitive system, it may be
possible to tie together seemingly unrelated phenomena
in experiments with normal subjects, as well as other
effects in brain-injured populations. We are currently
attempting to extend this account to the phenomena of
semantic satiation, verbal transformation, and priming (semantic
and repetition) in normal subjects, and perseverative naming
errors in aphasic patients. If successful, the theory should
be able to make testable predictions in neuroimaging
experiments, as well as in behavioral studies.
References
Byrne, J.H. (1982). Analysis of the synaptic depression contributing
to habituation of gill-withdrawal reflex in Aplysia californica. Journal
of Neurophysiology, 48, 431-438.
Gingrich, K.J. & Byrne, J.H. (1985). Simulation of synaptic depression,
posttetanic potentiation, and presynaptic facilitation of synaptic potentials
from sensory neurons mediating gill-withdrawal reflex in Aplysia. Journal
of Neurophysiology, 53, 652-669.
Miller, E.K., Gochin, P.M., & Gross, C.G. (1991). Habituation-like decrease
in the responses of neurons in inferior temporal cortex of the macaque.
Visual Neuroscience, 7, 357-362.
Pearlmutter, B.A. (1989). Learning state space trajectories in recurrent neural
networks. Neural Computation, 1, 263-269.
Rapp, B. & Caramazza, A. (1993). On the distinction between deficits of
access and deficits of storage: A question of theory. Cognitive
Neuropsychology, 10, 113-141.
Staddon, J.E.R. & Higa, J.J. (1996). Multiple time scales in simple
habituation. Psychological Review, 103, 720-733.
Warrington, E.K. (1975). The selective impairment of semantic memory.
QJEP, 27, 635-657.
Warrington, E.K. & Cipolotti, L. (1996). Word comprehension: The
distinction between refractory and storage impairments. Brain,
119, 611-625.
Warrington, E.K. & McCarthy, R.A. (1983). Category specific access
dysphasia. Brain, 106, 859-878.
Warrington, E.K. & McCarthy, R.A. (1987). Categories of knowledge.
Brain, 110, 1273-1296.
Warrington, E.K. & Shallice, T. (1979). Semantic access dyslexia.
Brain, 102, 43-63.
Back to Steve's Home Page
This page last updated 14 December 1998.